
Virulence of the phage-resistant mutant strain
抗噬菌体突变菌株的毒性
The third step of the pipeline concludes with the virulence of the phage-resistant mutant strain. Phage-bacterial co-incubation was used to isolate an evolved phage-resistant mutant of PA4.6C, which identify as PA4.6C-R. EOP could not be calculated because no plaques were obtained on PA4.6C-R at 1e12 PFU/mL, which lead to no bacterial growth inhibition when adding ΦSB in different concentrations (Fig. 3E). Biofilm formation was not inhibited by the addition of ΦSB (1e9 PFU/mL) to PA4.6C-R (Supplementary Fig. S7A and B), which confirms PA4.6C-R resistance to ΦSB.
第三步是对抗性突变株的毒力进行研究。噬菌体-细菌共孵育用于分离出 PA4.6C 的进化抗噬菌体突变体,即 PA4.6C-R。由于在 1e12 PFU/mL 的浓度下 PA4.6C-R 没有获得斑块,因此无法计算 EOP,这导致在添加不同浓度的 ΦSB 时没有抑制细菌生长(图 3E)。在 PA4.6C-R 上添加 ΦSB (1e9 PFU/mL)不会抑制生物膜的形成(补充图 S7A 和 B),这证实了 PA4.6C-R 对 ΦSB 的抗性。
Genomic comparison between PA4.6C and PA4.6C-R (Supplementary Table S3) revealed 13 non-synonymous point mutations potentially affecting virulence: four point mutations were identified in the type-IV pili (TIVP) pilQ gene, which is important for TIVP biogenesis by building a secretin channel for pili extrusion63; one point mutation was identified in the TIVP prepilin peptidase/methyltransferase PilD; eight point mutations were identified in the gene for fucose‑binding lectin LecB, an outer membrane protein potentially involved in pathogenicity65,66.
PA4.6C 和 PA4.6C-R(补充表 S3)发现了 13 个可能影响毒力的非同义点突变:在第四型纤毛虫(TIVP)pilQ 基因中发现了 4 个点突变,该基因通过建立纤毛虫挤出的分泌素通道对 TIVP 的生物发生非常重要 63 ;在 TIVP 前纤毛虫肽酶/甲基转移酶 PilD 中发现了 1 个点突变;在岩藻糖结合凝集素 LecB 基因中发现了 8 个点突变,该基因是一种可能参与致病性的外膜蛋白 65,66 。
The growth rate of PA4.6C-R was 40.1% compared to PA4.6C (0.50 ± 0.06 for PA4.6C and 0.32 ± 0.01 for PA4.6CR, p < 0.005, Fig. 4A). The area under the growth curve (AUC) decreased by 22.0% (79,662.4 ± 3100.3 for PA4.6C and 62,168.9 ± 863.9 for PA4.6C-R, p < 0.005, Fig. 4A). Biofilm formation was reduced in PA4.6C-R compared to the ancestral strain (Supplementary Fig. S8A,B) but no differences were found in antibiotic susceptibility testing between strains (Supplementary Fig. S9A,B). THP-1 macrophages expressed similar amounts of IL-8 after stimulation with supernatants from PA4.6C and PA4.6C-R, which were both in the range of the negative controls (Supplementary Fig. S10), and there were no differences in LDH (data not shown), a marker of cellular toxicity.
与 PA4.6C 相比,PA4.6C-R 的增长率为 40.1%(PA4.6C 为 0.50 ± 0.06,PA4.6CR 为 0.32 ± 0.01,p < 0.005,图 4A)。生长曲线下面积(AUC)减少了 22.0%(PA4.6C 为 79,662.4 ± 3100.3,PA4.6C-R 为 62,168.9 ± 863.9,p < 0.005,图 4A)。与祖先菌株相比,PA4.6C-R 的生物膜形成减少了(补充图 S8A,B),但在菌株间的抗生素敏感性测试中未发现差异(补充图 S9A,B)。用 PA4.6C 和 PA4.6C-R 的上清液刺激 THP-1 巨噬细胞后,IL-8 的表达量相似,都在阴性对照的范围内(补充图 S10),细胞毒性标志物 LDH 也没有差异(数据未显示)。
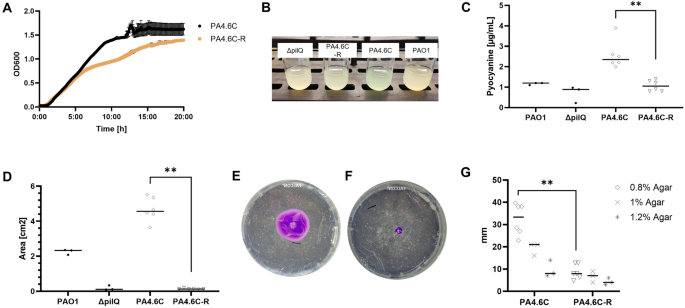
The following experiments were conducted as an attempt to account for changes in growth rate (normalization to 1e8 CFU/mL for 1:100 dilution of OD600 of 0.8 for PA4.6C and 1.0 for PA4.6C-R). Liquid cultures of PA4.6C were visually more blue-green compared to PA4.6C-R. Pyocyanin, a redox-active phenazine pigment responsible for the blue-green color of PSA, is involved in quorum sensing and virulence67. Quantification of pyocyanin revealed a decrease in pyocyanin production (58.4% decrease on average comparing PA4.6C to PA4.6C-R, p < 0.005, Fig. 4B,C). Elastase, an extracellular protease of PSA, has been described to facilitate tissue destruction and thereby host invasion68,69. Elastase production was low in both the ancestral and mutant strains (both < 0.079U/mL). Twitching motility assays presented a clearly visible, and quantifiable, decrease in motility for the mutant strain PA4.6C-R in the standard twitching motility assay (p < 0.005), and for the adapted twitching motility assay (0.8% agar, p < 0.005, see Fig. 4D-G). Swimming and swarming motility were not significantly different between PA4.6C and PA4.6C-R (Supplementary Fig. S11A,B).
以下实验试图解释生长率的变化(PA4.6C 的 OD 600 为 0.8,PA4.6C-R 的 OD 600 为 1:100 稀释,归一化为 1e8 CFU/mL)。与 PA4.6C-R 相比,PA4.6C 的液体培养物在视觉上呈蓝绿色。焦花青素是一种具有氧化还原作用的酚嗪色素,是造成 PSA 呈蓝绿色的原因,它参与了法定人数感应和毒力 67 。对焦花青素的定量分析显示,PSA 的焦花青素产量有所减少(PA4.6C 与 PA4.6C-R 相比平均减少了 58.4%,p < 0.005,图 4B、C)。弹性蛋白酶是 PSA 的一种细胞外蛋白酶,已被描述为可促进组织破坏,从而促进宿主入侵 68,69 。祖代菌株和突变菌株的弹性蛋白酶产量都很低(均小于 0.079U/mL)。抽动运动试验显示,在标准抽动运动试验(p < 0.005)和适应性抽动运动试验(0.8% 琼脂,p < 0.005,见图 4D-G)中,突变株 PA4.6C-R 的运动能力明显下降,而且可以量化。PA4.6C 和 PA4.6C-R 的游泳和成群运动没有明显差异(补充图 S11A、B)。
Amplification 放大
The fourth step of the preparation pipeline for emergency phage therapy pertains to amplification. Amplification for the final phage preparation was performed with a total of 1L of ~ 0.01 MOI of the PVS with PAO1 in Tryptic Soy Broth of non-animal origin. Amplification and subsequent dialysis in PBS supplemented with 1 M MgSO4 resulted in a final, sterile-filtered, visually clear, and transparent 10 mL solution of ΦSB with a titer of 2e11 PFU/mL, which was found to be stable for at least 3 months at 4 °C. This solution was thus ready for dilution to the appropriate titer and buffer (e.g., saline for intravenous administration, PBS for inhalation, or for preparation with a formulation such as an ointment with an appropriate pH for respective applications). This sample would be sent for external endotoxin testing and USP < 71 > sterility for potential use for phage therapy (data not shown).
应急噬菌体疗法制备流程的第四步是扩增。最终噬菌体制备的扩增是在非动物源胰蛋白酶大豆肉汤中用 1 升约 0.01 MOI 的 PVS 与 PAO1 进行的。放大后,在添加了 1 M MgSO 4 的 PBS 中进行透析,最终得到无菌过滤、清澈透明的 10 mL ΦSB 溶液,滴度为 2e11 PFU/mL。因此,该溶液可稀释至适当的滴度和缓冲液(例如,静脉注射用生理盐水、吸入用 PBS,或配制成具有适当 pH 值的软膏等制剂)。该样本将送去进行外部内毒素检测和 USP < 71 > 无菌检测,以便用于噬菌体治疗(数据未显示)。